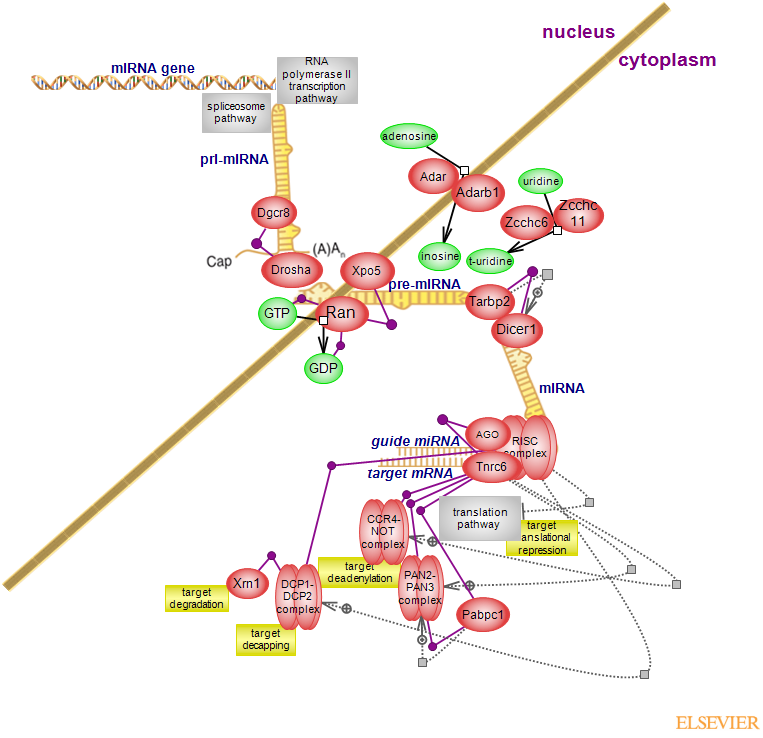
Small non-coding RNAs have emerged as potent regulators of gene expression. Of the three classes - microRNA (miRNA), endogenous small interfering RNA (endo-siRNA) and Piwi-interacting RNA (piRNA), miRNA is the better understood system. Its main function is post-transcriptional repression of target genes. miRNAs are transcribed from individual genes or from the introns of intron-containing genes. The resulting primary miRNA transcript (pri-miRNA) contains a hairpin-loop structure with single str
anded arms flanking the hairpin. Many have a cap and a polyA tail indicative of polymerase II transcription. The majority of miRNA are products of pol II transcription; less frequently, of pol III. In the nucleus, the pri-miRNA is cleaved to a ~60-70 nucleotides stem-looped precursor (pre-miRNA) by the microprocessor complex. The core components of the complex are the RNase III Drosha (Rnasen) and its cofactor Dgcr8, essential for determining the cleavage site. The two RNase domains of the enzyme cleave the 5¿ and 3¿ single strands about 11 bp away from the junction of single- and double-stranded RNA. In a Drosha-independent manner, miRNAs can be generated from pre-miRNA-like introns, called mirtrons; the Drosha-mediated route is the canonical miRNA pathway. pre-miRNA is recognized by Xpo5 - a member of the nuclear transport receptor family which, in conjunction with GTP-bound Ran, transports it into the cytoplasm. The HEAT repeats of Xpo5 are important for both pre-miRNA and GTP-Ran binding. Hydrolysis of GTP to GDP in the cytoplasm leads to release of cargo. In the cytosol, pre-miRNA is processed by Dicer1 - the other RNase III in the pathway. Dicer1, aided by Tarbp2 (Trbp), cleaves the hairpin-loop of pre-miRNA to produce a ~22 nucleotide duplex. Binding of Tarbp2 to the helicase-like domain of Dicer induces conformational changes that abrogate its inhibitory effect. The relative thermodynamic stability of the base pairs at the ends of the duplex determines which strand will become the mature or guide strand. The strand with the less stable 5'end becomes the guide of the miRNA-induced silencing complex (RISC) - the effector complex of miRNA pathway. Members of the argonaute family of proteins are core components of RISC complex; its assembly however, is not well understood and appears to vary between species. In mammals, four proteins of the AGO subfamily with redundant functions are involved in miRNA and siRNA pathways. The ~100kDa proteins contain characteristic PAZ, MID (middle) and PIWI domains; PAZ and MID contact the 3' and 5' end of miRNA, respectively. The C-terminal PIWI domain may have endonucleolytic activity, while the N-terminal domain facilitates duplex unwinding. Drosha and Dicer1 generate varied length products; these other length variants, called isomiRs may vary in their gene expression regulation or may regulate the same targets as the canonical miRNAs.
miRNAs and target mRNAs form partially complementary duplexes with the 5' 2-7 nucleotide 'seed' sequence of miRNA and the 3' UTR sequence of target mRNA. Although the 3'UTR was believed to be the major miRNA binding site, recent results indicate that miRNA binding sites may be located within the 5'UTR and the coding sequence (CDS). The translational repression of target genes may occur at the initiation and post initiation steps; the molecular mechanisms are poorly understood. miRNA can also mediate destabilization of mRNA by inducing its deadenylation, leading to decapping and degradation. Tnrc6 (GW182) proteins - the second component of RISC acting downstream of AGO, contact AGOs via the N-terminal GW repeats; the glycine-tryptophan or tryptophan-glycine repeats (GW/WG) are a characteristic feature of the proteins. The N-terminal repeats contain multiple, independent binding sites for AGO referred to as AGO hooks. N- and C-terminal repeats interact with Cnot1 and C-terminal repeats and PAM2 motif interact with Pan3 - subunits of CCR4-NOT and PAN2-PAN3 deadenylase complexes, respectively. The C-terminal region of fly and mammalian proteins also contains a glutamine (Q)-rich domain, a ubiquitin binding domain (UBA), the PAM2 motif and the non-canonical RNA recognition motif (RRM). PAM2 motif also interacts with the poly A binding protein Pabpc1. The PAM2 motif of Pan3 binds Pabpc1, which stimulates the catalytic activity of the complex. Deadenylation is accompanied by decapping via Dcp1-Dcp2 complex, followed by Xrn1 mediated degradation. Dcp2 catalyzes the hydrolysis of the 5' cap structure, Dcp1 is its main activator. Additional activators/decapping factors are present, of which some are species specific. The association of a subset of these factors with miRNA targets is promoted by RISC via both AGO and Tnrc6 mediated interactions. Editing of RNA molecule can serve as a control and regulate the pathway: deamination of adenosine to inosine can affect processing by either RNase processing or, in some cases, result in differential targeting; 3' uridylation can inhibit or promote the activity of Dicer but largely, has an inhibitory effect. Of the three deaminases, Adar (Adar1) and Adarb1 (Adar2) are ubiquitously expressed and have multiple isoforms, of which some shuttle between the nucleus and cytoplasm. Uridylation is carried out by two cytoplasmic uridyl transferases: Zcchc6 (TUT7) and Zcchc11 (TUT4). In addition, miRNAs appear to be involved in the control of their own biogenesis. The molecular mechanisms of these regulatory layers remain to be elucidated, Aberrant expression of miRNAs has been observed for many conditions including various types of cancer.To see the ontology report for annotations, GViewer and download click here...(less)