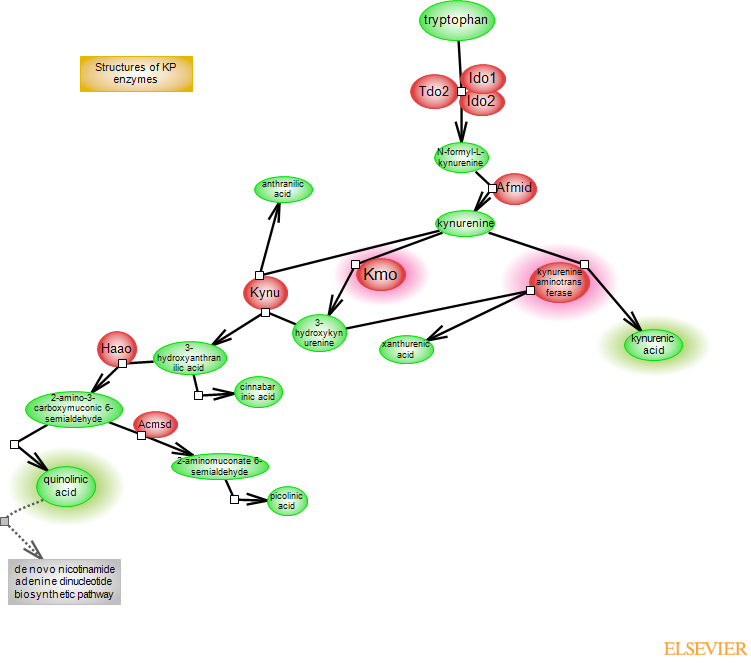
Kynurenine, the central component of the kynurenine metabolic pathway, is a product of tryptophan catabolism. Tryptophan also serves as the precursor for the production of serotonin, which in turn, serves as a precursor to melatonin. Most of tryptophan (~95%) is channeled into the kynurenine pathway. The incorporation of oxygen into the indole moiety of tryptophan in the first and rate-limiting step of kynurenine metabolism is catalyzed by three heme-containing enzymes in mammals: tryptophan-2,
3-dioxygenase (TDO2) and indoleamine-2,3 dioxygenase 1 and 2 (IDO1 and IDO2). The product of the reaction is N-formylkynurenine, converted to kynurenine (KYN) by arylformamidase (Afmid). KYN is a branching point for two metabolic routes. In one, it is converted to kynurenic acid (KYNA) by kynurenine aminotransferases (KATs). In the other, it initiates the production of quinolinic acid (QUIN). Here, KYN is hydroxylated to 3-hydroxykynurenine (3-HK) by kynurenine 3-monooxygenase (KMO). 3-HK is cleaved by kynureninase (KYNU) to 3-hydroxyanthranilic acid (3-HAA), which is then oxidized by 3-HAA oxygenase (3-HAO/HAAO) to 2-amino-3-carboxymuconic 6-semialdehyde (ACMS). ACMS spontaneously rearranges to quinolinic acid (QUIN). In the brain, the two routes are separated: KATs are primarily expressed in astrocytes whereas KMO is expressed in microglia. KYNA is an antagonist of NMDA and alpha7 nicotinic acetylcholine receptors, and may act as a radical scavenger. QUIN is the precursor of nicotinamide adenine dinucleotide (NAD+) coenzyme in the de novo NAD+ biosynthetic pathway. NAD+ is a widely-used coenzyme involved in redox reactions and is also used as a substrate by enzymes involved in posttranslational modifications. QUIN is a potentially neurotoxic metabolite; it selectively activates NMDA receptors and it can promote the production of reactive oxygen species (ROS). Other KYN metabolites include anthranilic acid (AA), metabolized from KYN by KYNU or xanthurenic acid (XA), metabolized from 3-HK by KATs. XA may have a role in hippocampal function. Cinnabarinic acid (CA) results from auto-oxidation of 3-HAA and could act as a partial agonist of metabotropic glutamate receptor 4. ACMS can be metabolized by ACMS decarboxylase (ACMSD) to 2-aminomuconate 6-semialdehyde (AMS), which rearranges to picolinic acid (PA). However, the expression of Acmsd is low and PA is not a major route of ACMS metabolism. Deregulation of the kynurenine pathway is associated with several neurological and neurodegenerative conditions. Neuroinflammation and microglia activation with increases in KMO activity and QUIN-dependent excitotoxicity have been associated with the pathogenesis of Huntington's disease (HD); elevated QUIN levels are also observed in other neurodegenerative conditions. On the other hand, despite its overall neuroprotective effects, elevated KYNA, due to its antagonistic impact on NMDA and cholinergic systems, can be detrimental. KMO polymorphism, decreased KMO expression and increased KYNA levels have been associated with bipolar disorder and schizophrenia. Enzymes in the kynurenine pathway are targets for drug development.
Enzymes and reaction steps
Tdo2, Ido1 and Ido2 are the enzymes of the first reaction step in the kynurenine metabolic pathway. Tdo2 is a tetrameric protein with specificity for L-tryptophan, whereas the IDO enzymes are monomeric with a wider range of substrates. Tdo2 is constitutively expressed in liver and at a lower level in neurons, astrocytes and endothelial cells. The human gene contains 12 exons and 11 introns; the protein tetramer is a dimer of dimers, and each monomer contains a heme binding site that can be in an opened or closed conformation. The catalysis of L-tryptophan to N-formylkynurenine by Tdo2 involves incorporation of both atoms of oxygen molecules into the indole moiety. Activation of Tdo2 and subsequent production of kynurenine, which can cross the blood brain barrier and promote the local production of its metabolites, is believed to underlie the involvement of Tdo2 in brain diseases. Tdo2 is a target for the development of inhibitory drugs. Ido1 is expressed in several different cell types including neurons, astrocytes and microglia; expression can be constitutive but it is strongly induced by inflammatory and immune stimuli. Ido2 expression is more restricted and relatively less is known about this particular enzyme. Reaction mechanisms, heme-coordinating residues and enzyme cofactors have been hypothesized or proposed. Crystal structures for TDO2 and IDO1 are available.
The hydrolysis of N-formylkynurenine to kynurenine (KYN) is carried out by arylformamidase (Afmid). Modeling and mutagenesis studies in mouse predict a catalytic triad formed by serine, aspartic acid and histidine residues (S162, D247, H279, mouse notation).
The irreversible transamination of KYN to KYNA is carried out by kynurenine aminotransferases (KATs). The four enzymes belong to the alpha family of pyridoxal 5'-phosphate (PLP)-dependent aminotransferases and are assigned to fold type I. KYAT1 (KatI) and KYAT3 (KatIII) share the highest sequence identity. GOT2 (KatIV) and AADAT (KatII) are mitochondrial proteins. The four enzymes transaminate KYN as well as other substrates. KYN transamination occurs in two steps: the first step, mediated by the Kat enzymes, produces an unstable ketoacid intermediate which rapidly undergoes an intramolecular cyclization to KYNA. There are differences in substrate specificity and responses to inhibitors between the four enzymes, and their exact contributions to KYNA synthesis in the brain remain to be assessed. Crystal structures for the four proteins, with or without bound substrate, are available. The overall similar structures reveal a homodimer with two active sites at the interface between subunits but also point to subtle differences and different conformational changes that occur upon substrate binding.
The formation of quinolinic acid (QUIN) is initiated by the kynurenine 3-monooxygenase (KMO), a flavin adenine dinucleotide (FAD)-containing enzyme in the outer mitochondrial membrane. FAD is bound non-covalently but tightly to the enzyme, and catalysis requires the reduction of FAD by NADH or NADPH, the latter being preferred. The product of the KMO-catalyzed reaction is 3-HK. Structures of C-terminal truncated protein from yeast, which unlike truncated proteins from other species is enzymatically active, with or without a bound inhibitor, are available.
Kynureninase (KYNU) catalyzes the hydrolysis of 3-HK to 3-HAA. As a member of the aminotransferase superfamily, Kynu uses PLP as a cofactor and acts as a dimer. 3-HAA in turn is a substrate for HAAO, a non-heme iron-dependent enzyme that catalyzes cleavage of the benzene ring of 3-HAA to produce 2-amino-3-carvoxymuconic 6-semialdehyde, which spontaneously rearranges to QUIN.
The important metabolites - KYNA and QUIN, and the branching point enzymes leading to their production - KATs and KMO, are highlighted.