|
|

|
CHROMATIN REMODELING PATHWAY (PW:0001339)
Description
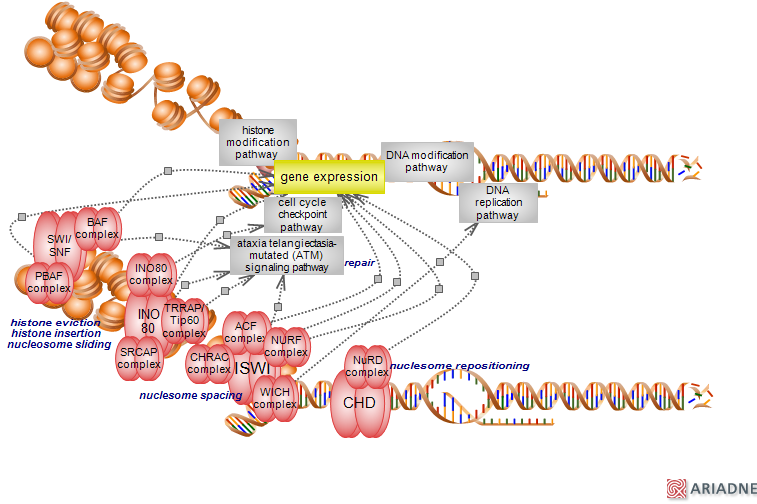
Epigenetic changes, the modification and remodeling of chromatin, play a key role in the differential expression of genes. Chromatin modification and remodeling directly affect the relative relaxation or compaction of chromatin and thus, the extent to which DNA replication, transcription, damage response and repair, associated RNA processing and splicing are promoted or are silenced, respectively. The basic unit of chromatin is the nucleosome - it consists of 145-147 base pairs of DNA wrapped around an octameric structure formed by core histones. Repeating nucleosomes assemble into higher-order structures stabilized by a linker histone. Chromatin modification involves the methylation of cytosine residues in DNA and the many ways in which the histone residues are modified. The remodeling of chromatin involves the ATP-dependent sliding, spacing and repositioning of entire nucleosomes, insertion, eviction or exchange of histones, chromatin assembly and disassembly. It is carried out by four major protein complexes representing four remodeling pathways with distinct functions. Epigenetic changes are heritable and independent of DNA sequence. However, DNA sequence variation can affect the function of players involved and thus, of epigenetic outcomes. An intimate dialog exists between the pathways of chromatin modification and remodeling to finely tune the epigenetic responses. The four remodeling pathways represent the subdivision of catalytic subunits of chromatin remodelers - members of the DEAD/H family of ATP-binding helicases, into four families whose function and associated complexes perform specific, non-redundant functions. The four families/complexes are: SWI/SNF, INO80(SWRI), ISWI and CHD. Chromatin remodeling complexes (CRCs) interact with nucleosomal DNA and histones, with modified histones and with other proteins via several DNA and protein recognition domains. The four remodeling complexes are individually described.
SWI/SNF complex
SWI/SNF complex, originally identified in yeast, was subsequently identified in fly and mammals. There are several mammalian SWI/SNF complexes of which the main ones are BAF (yeast SWI/SNF-A) and PBAF (yeast SWI/SNF-B), present in the same cell type (shown). Others are cell-specific and represented by neural progenitor BAF (npBAF), neuron BAF (nBAF) and embryonic stem cell BAF (esBAF) (not shown). All complexes contain one of the two catalytic subunits Smarca4/Brg1 or Smarca2/Brm, share several other core subunits and include accessory subunits specific to given complexes. The presence of subunits is in many cases, mutually exclusive. Many subunits are represented by gene families allowing for the combinatorial assembly of complexes and are subject to gene expression and developmental programs. Subunit exchange is also a feature of SWI/SNF assembly, further increasing the number of possible complex instances, estimated to be in excess of seventy. Besides the catalytic subunits, the other core subunits include Smarcc1/Baf155 or Smarcc2/Baf170, Smarcd1, 2 or 3 (Baf60a/b/c), Actl6A or B (Baf53a/b) and Smarcb1 (Baf47/Snf5). Additional subunits include Arid1A or B (Baf250a/b ) or Arid2 (Baf200), Pbrm1 (Baf180), Phf10, Dpf1, 3 or 2 (Baf45a/b/c/d), Brd7, Brd9, Smarce1 (Baf57) and beta-actin. The two catalytic subunits - Smarca4 and 2 have N-terminal DEXD and HELIC subdomains and C-terminal bromodomains that can bind acetylated histone tails. They have ATPase and translocase activity but no helicase activity. Other domains present in the various subunits of the complexes facilitate the interaction with DNA ARID, HMG zinc finger, leucine zipper domains; or the interaction with proteins - LXXLL, BAH, SANT, SWIRM, SWIB domains. The mammalian SWI/SNF complexes be found within intergenic regions and also promoters. They can slide nucleosomes, eject and insert histone octamers, are involved in chromatin opening, chromosome condensation and segregation and exert both activation and repression of genes. They also play a role in the DNA repair. The structure of an yeast complex related to ySWI/SNF, show the nucleosome being engulfed within a central cavity. Mutations in several subunits of BAF and PBAF complexes are present in 20% of human cancers and they include both catalytic subunits as well as shared and unique components. Most are loss of functions resulting from frameshift, deletion and nonsense. To see the ontology report for annotations, GViewer and download, click here
INO80 complex
The INO80 remodeler has been identified in yeast, fly and mammals and is present as several complexes; the mammalian complexes are INO80, SRCAP (yeast SWRI or SWR-C) and TRAAP/Tip60 (yeast NuA4). Unlike the other remodeling factors, the INO80 catalytic subunits have helicase activity. A unique features of the enzymes is the presence of an insertion that creates a split ATPase domain which allows for the recruitment of Ruvbl1 and 2 (Tip49a and b) helicases - hexameric AAA+ ATPases related to the microbial RuvB helicase. They are present in a 6:1 stoichiometry suggestive of a hexameric structure, similar to the bacterial helicase. Structural studies indicate a rather compact topology for the SWR-C complex whereas the INO80 displays an elongated shape with the more striking feature involving the RuvB module. The INO80 structure harbors a double-ring heterododecamer, whereas the SWR-C complex has a single-ring heterohexamer. The nucleosome binding mode reveals a 'clinging' feature for the more compact SWR-C complex, while the elongated INO80 appears to 'grasp' the nucleosome. In addition to the ATPase domain, the enzymes also contain a conserved helicase-SANT-associated (HAS) domain that may be involved in the interaction with actin-related proteins (ACTRs/ARPs). The Ep400 ATPase of the TRAAP/Tip60 complex also contains a carboy-terminal SANT domain that may facilitate the interaction with histones. In addition the the Ruvbl helicases, other components include members of ACTRs/ARPs such as Actr4 present in all complexes, beta-actin and Actl6a (Baf53a). Other subunits are unique or shared by a subset of complexes. The complexes can slide nucleosomes and possibly evict histones and they have an affinity for histone variants, in particular for the H2A variants H2AZ and H2AX. A special feature of the INO80 complexes is their role in processes other than regulation of transcription with demonstrated roles in the double strand DNA repair downstream of ATM signaling and cell cycle checkpoint - functions dependent on their ability to incorporate the H2A.Z histone variant. To see the ontology report for annotations, GViewer and download, click here
ISWI complex
The ISWI complex has first been identified in Drosophila with orthologs in yeast and mammalian systems. Like the other remodelers, ISWI is present as several complexes which in humans are primarily represented by ACF, CHRAC and NURF. WICH is an additional ISWI complex present in mammals with a specific role in the replication of DNA in condensed regions. THE ISWI members of the DEAD/H-related enzymes, in addition to the ATPase domain located in the N-terminal part of the protein, contain HAND, SANT and SLIDE domains within the C-terminal part of the protein. The stoichiometry of ISWI to nucleosome is reported to be 2:1. ACF, CHRAC and WICH share the same catalytic subunit Smarca5/Snf2h. The non-catalytic Baz1a (Wcrf180/Acf1) is shared by ACF and CHRAC with CHRAC having two additional unique subunits - Chrac1 (Chrac15) and Pole3 (Chrac17). WICH has the non-catalytic Baz1b (Wstf) as a component. NURF complex has its own members and is composed of Smarca1 (Snf2l), Bptf, Rbbp7 (RbAp46) and Rbb4 (RbAp48). ISWI complexes are involved in nucleosome spacing which facilitates the formation of chromatin leading to gene silencing. They can also switch nucleosome to clear or block promoters, thus promoting both and activating functions in the regulation of gene expression. In addition to the regulation of DNA replication and of transcription by all three RNA polymerases, ISWI complexes are also involved in DNA repair. To see the ontology report for annotations, GViewer and download, click here
CHD complex
The CHD family of ATPases, in addition to the conserved DEAD/H ATPase domain, the have N-terminal tandem chromodomains. The nine members of the family have been subdivided into three families based on the domain structure. Of these, Chd3 and 4 are components of a large complex that has both chromatin remodeling and histone deacetylase activities, known as NuRD. Chd5 family member appears to be preferentially, though not exclusively expressed in the central nervous system and testis. The three ATPases also have two plant homeodomain (PHD) fingers and two domains of unknown function (DUF) which despite their name, show repressive transcriptional activity via association with corepressors. The deacetylase members of NuRD are Hdac1 and 2. Non-enzymatic components include the Mbd2 and 3 proteins with methyl CpG binding domains, the Mta family members 1, 2 and 3, the zinc finger proteins Gatad2a and 2b (p66alpha and beta) and the retinoblastoma binding proteins4 and 7. Several components also establish interactions with coregulators. NuRD - originally identified as silencer of gene expression, appears to fine-tune transcription and functions in many contexts. Other regulatory roles include DNA repair, genome integrity and cell cycle progression. NuRD directly controls the levels of acetylated p53 and therefore indirectly, cell cycle checkpoint. Chd4 appears to also have NuRD independent functions, such as its role in DNA damage repair. Whether this also implicates NuRD, remains to be established. Alterations of this remodeling pathway have been associated with developmental defects, aging and oncogenesis. Interestingly, NuRD appears to have opposing effects in cancer acting both as an oncogene and as a tumor suppressor. To see the ontology report for annotations, GViewer and download, click here ...(less) |
Pathway Diagram:
|
|
|
Genes in Pathway:
|
|
| G
|
Actl6a
|
actin-like 6A
|
|
ISO
|
|
RGD |
PMID:23355908 |
RGD:8694154 |
NCBI chr 2:117,413,206...117,436,758
Ensembl chr 2:117,418,601...117,436,754
|

|
| G
|
Arid1a
|
AT-rich interaction domain 1A
|
|
ISO
|
|
RGD |
PMID:23355908 |
RGD:8694154 |
NCBI chr 5:151,192,014...151,269,291
Ensembl chr 5:151,192,014...151,265,386
|
|
| G
|
Arid1b
|
AT-rich interaction domain 1B
|
|
ISO
|
|
RGD |
PMID:23355908 |
RGD:8694154 |
NCBI chr 1:47,973,199...48,328,793
Ensembl chr 1:47,973,997...48,328,793
|
|
| G
|
Pbrm1
|
polybromo 1
|
|
ISO
|
|
RGD |
PMID:23355908 |
RGD:8694154 |
NCBI chr16:6,219,933...6,319,336
Ensembl chr16:6,219,917...6,319,336
|
|
| G
|
Smarca4
|
SWI/SNF related BAF chromatin remodeling complex subunit ATPase 4
|
|
ISO
|
|
RGD |
PMID:23355908 |
RGD:8694154 |
NCBI chr 8:28,438,370...28,535,071
Ensembl chr 8:28,443,810...28,535,070
|
|
|
|
| G
|
Chd3
|
chromodomain helicase DNA binding protein 3
|
|
ISO
|
|
RGD |
PMID:24880148 |
RGD:9585661 |
NCBI chr10:54,562,437...54,588,842
Ensembl chr10:54,562,437...54,588,810
|
|
| G
|
Chd4
|
chromodomain helicase DNA binding protein 4
|
|
ISO
|
|
RGD |
PMID:24880148 |
RGD:9585661 |
NCBI chr 4:159,584,623...159,617,867
Ensembl chr 4:159,584,654...159,617,866
|
|
| G
|
Chd5
|
chromodomain helicase DNA binding protein 5
|
|
ISO
|
|
RGD |
PMID:24880148 |
RGD:9585661 |
NCBI chr 5:168,130,851...168,181,705
Ensembl chr 5:168,130,575...168,179,441
|
|
| G
|
Gatad2a
|
GATA zinc finger domain containing 2A
|
|
ISO
|
|
RGD |
PMID:24880148 |
RGD:9585661 |
NCBI chr16:19,463,500...19,553,497
Ensembl chr16:19,463,500...19,553,497
|
|
| G
|
Gatad2b
|
GATA zinc finger domain containing 2B
|
|
ISO
|
|
RGD |
PMID:24880148 |
RGD:9585661 |
NCBI chr 2:178,046,192...178,127,633
Ensembl chr 2:178,046,085...178,147,315
|
|
| G
|
Hdac1
|
histone deacetylase 1
|
|
ISO
|
|
RGD |
PMID:24880148 |
RGD:9585661 |
NCBI chr 5:147,138,328...147,165,387
Ensembl chr 5:147,138,156...147,165,387
|
|
| G
|
Hdac2
|
histone deacetylase 2
|
|
ISO
|
|
RGD |
PMID:24880148 |
RGD:9585661 |
NCBI chr20:42,101,815...42,126,486
Ensembl chr20:42,102,861...42,126,311
|
|
| G
|
Mbd2
|
methyl-CpG binding domain protein 2
|
|
ISO
|
|
RGD |
PMID:24880148 |
RGD:9585661 |
NCBI chr18:66,449,196...66,516,112
Ensembl chr18:66,449,358...66,516,111
|
|
| G
|
Mbd3
|
methyl-CpG binding domain protein 3
|
|
ISO
|
|
RGD |
PMID:24880148 |
RGD:9585661 |
NCBI chr 7:9,961,053...9,970,845
Ensembl chr 7:9,963,797...9,970,265
|
|
| G
|
Mta1
|
metastasis associated 1
|
|
ISO
|
|
RGD |
PMID:24880148 |
RGD:9585661 |
NCBI chr 6:137,999,845...138,038,696
Ensembl chr 6:137,999,915...138,038,696
|
|
| G
|
Mta2
|
metastasis associated 1 family, member 2
|
|
ISO
|
|
RGD |
PMID:24880148 |
RGD:9585661 |
NCBI chr 1:215,267,286...215,276,008
Ensembl chr 1:215,267,046...215,276,062
|
|
| G
|
Mta3
|
metastasis associated 1 family, member 3
|
|
ISO
|
|
RGD |
PMID:24880148 |
RGD:9585661 |
NCBI chr 6:16,619,575...16,739,732
Ensembl chr 6:16,621,998...16,739,485
|
|
| G
|
Rbbp4
|
RB binding protein 4, chromatin remodeling factor
|
|
ISO
|
|
RGD |
PMID:24880148 |
RGD:9585661 |
NCBI chr 5:146,940,207...146,959,501
Ensembl chr 5:146,922,611...146,959,501
|
|
| G
|
Rbbp7
|
RB binding protein 7, chromatin remodeling factor
|
|
ISO
|
|
RGD |
PMID:24880148 |
RGD:9585661 |
NCBI chr X:35,544,873...35,563,030
Ensembl chr X:35,544,873...35,563,030
|
|
|
|
| G
|
Actb
|
actin, beta
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr12:16,776,664...16,779,634
Ensembl chr12:16,776,661...16,780,242
|
|
| G
|
Actl6a
|
actin-like 6A
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 2:117,413,206...117,436,758
Ensembl chr 2:117,418,601...117,436,754
|
|
| G
|
Actr5
|
actin related protein 5
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 3:167,702,299...167,715,736
Ensembl chr 3:167,702,482...167,715,730
|
|
| G
|
Actr6
|
actin related protein 6
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 7:26,037,574...26,058,362
Ensembl chr 7:26,037,574...26,060,604
|
|
| G
|
Actr8
|
actin related protein 8
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr16:5,170,649...5,192,883
Ensembl chr16:5,167,237...5,186,180
|
|
| G
|
Brd8
|
bromodomain containing 8
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr18:26,455,824...26,504,106
Ensembl chr18:26,455,512...26,504,006
|
|
| G
|
Dmap1
|
DNA methyltransferase 1-associated protein 1
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 5:136,380,211...136,388,229
Ensembl chr 5:136,380,216...136,388,219
|
|
| G
|
Ep400
|
E1A binding protein p400
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr12:51,551,665...51,658,635
Ensembl chr12:51,551,695...51,658,634
|
|
| G
|
Epc1
|
enhancer of polycomb 1
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr17:59,198,513...59,289,535
Ensembl chr17:59,197,677...59,289,519
|
|
| G
|
Epc2
|
enhancer of polycomb 2
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 3:54,276,187...54,376,373
Ensembl chr 3:54,276,239...54,376,916
|
|
| G
|
Ing3
|
inhibitor of growth family, member 3
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 4:51,439,218...51,469,502
Ensembl chr 4:51,443,763...51,470,951
|
|
| G
|
Ino80
|
INO80 complex ATPase subunit
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 3:126,822,280...126,919,532
Ensembl chr 3:126,822,280...126,919,532
|
|
| G
|
Ino80b
|
INO80 complex subunit B
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 4:117,185,427...117,188,930
Ensembl chr 4:117,185,620...117,189,168
|
|
| G
|
Ino80c
|
INO80 complex subunit C
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr18:15,653,508...15,752,455
Ensembl chr18:15,653,511...15,668,144
|
|
| G
|
Ino80d
|
INO80 complex subunit D
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 9:71,940,941...72,007,879
Ensembl chr 9:71,951,031...72,040,232
|
|
| G
|
Ino80e
|
INO80 complex subunit E
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 1:190,891,315...190,902,902
Ensembl chr 1:190,891,936...190,902,541
|
|
| G
|
Kat5
|
lysine acetyltransferase 5
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 1:212,325,089...212,332,640
Ensembl chr 1:212,325,090...212,332,587
|
|
| G
|
Mcrs1
|
microspherule protein 1
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 7:132,262,719...132,271,533
Ensembl chr 7:132,262,722...132,271,533
|
|
| G
|
Meaf6
|
MYST/Esa1-associated factor 6
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 5:142,628,281...142,653,076
Ensembl chr 5:142,628,968...142,656,293
|
|
| G
|
Morf4l1
|
mortality factor 4 like 1
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 8:99,522,801...99,544,432
Ensembl chr 8:99,500,898...99,544,331
|
|
| G
|
Morf4l2
|
mortality factor 4 like 2
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr X:104,874,850...104,885,946
Ensembl chr X:104,852,693...104,892,855
|
|
| G
|
Mrgbp
|
MRG domain binding protein
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 3:188,074,695...188,078,162
Ensembl chr 3:188,074,709...188,078,153
|
|
| G
|
Nfrkb
|
nuclear factor related to kappa B binding protein
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 8:38,089,574...38,121,506
Ensembl chr 8:38,089,999...38,121,501
|
|
| G
|
Ruvbl1
|
RuvB-like AAA ATPase 1
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 4:122,489,754...122,524,666
Ensembl chr 4:122,489,680...122,524,665
|
|
| G
|
Ruvbl2
|
RuvB-like AAA ATPase 2
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 1:105,038,487...105,051,851
Ensembl chr 1:105,038,174...105,051,720
|
|
| G
|
Srcap
|
Snf2-related CREBBP activator protein
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 1:191,554,043...191,607,096
Ensembl chr 1:191,554,043...191,604,265
|
|
| G
|
Tfpt
|
TCF3 fusion partner
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 1:74,502,994...74,512,766
Ensembl chr 1:74,502,994...74,513,370
|
|
| G
|
Trrap
|
transformation/transcription domain-associated protein
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr12:14,851,758...14,941,444
Ensembl chr12:14,851,758...14,941,407
|
|
| G
|
Uchl5
|
ubiquitin C-terminal hydrolase L5
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr13:58,070,947...58,106,825
Ensembl chr13:58,042,029...58,107,441
|
|
| G
|
Vps72
|
vacuolar protein sorting 72 homolog
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 2:185,367,329...185,379,199
Ensembl chr 2:185,367,623...185,379,191
|
|
| G
|
Yeats4
|
YEATS domain containing 4
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 7:54,744,220...54,751,361
Ensembl chr 7:54,744,221...54,751,361
|
|
| G
|
Yy1
|
YY1 transcription factor
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr 6:133,471,615...133,500,875
Ensembl chr 6:133,471,562...133,497,257
|
|
| G
|
Znhit1
|
zinc finger, HIT-type containing 1
|
|
ISO
|
|
RGD |
PMID:21502417 |
RGD:9495926 |
NCBI chr12:25,323,292...25,329,566
Ensembl chr12:25,323,114...25,329,563
|
|
|
|
| G
|
Baz1a
|
bromodomain adjacent to zinc finger domain, 1A
|
|
ISO
|
|
RGD |
PMID:21358755 |
RGD:9495920 |
NCBI chr 6:78,124,872...78,247,672
Ensembl chr 6:78,124,872...78,247,648
|
|
| G
|
Baz1b
|
bromodomain adjacent to zinc finger domain, 1B
|
|
ISO
|
|
RGD |
PMID:21358755 |
RGD:9495920 |
NCBI chr12:27,068,541...27,126,511
Ensembl chr12:27,068,541...27,126,511
|
|
| G
|
Bptf
|
bromodomain PHD finger transcription factor
|
|
ISO
|
|
RGD |
PMID:21358755 |
RGD:9495920 |
NCBI chr10:92,480,007...92,582,485
Ensembl chr10:92,480,007...92,582,413
|
|
| G
|
Chrac1
|
chromatin accessibility complex subunit 1
|
|
ISO
|
|
RGD |
PMID:21358755 |
RGD:9495920 |
NCBI chr 7:106,901,841...106,905,441
|
|
| G
|
Pole3
|
DNA polymerase epsilon 3, accessory subunit
|
|
ISO
|
|
RGD |
PMID:21358755 |
RGD:9495920 |
NCBI chr 5:80,989,508...80,992,742
Ensembl chr 5:80,989,511...80,992,742
Ensembl chr 7:80,989,511...80,992,742
|
|
| G
|
Rbbp4
|
RB binding protein 4, chromatin remodeling factor
|
|
ISO
|
|
RGD |
PMID:21358755 |
RGD:9495920 |
NCBI chr 5:146,940,207...146,959,501
Ensembl chr 5:146,922,611...146,959,501
|
|
| G
|
Rbbp7
|
RB binding protein 7, chromatin remodeling factor
|
|
ISO
|
|
RGD |
PMID:21358755 |
RGD:9495920 |
NCBI chr X:35,544,873...35,563,030
Ensembl chr X:35,544,873...35,563,030
|
|
| G
|
Smarca1
|
SNF2 related chromatin remodeling ATPase 1
|
|
ISO
|
|
RGD |
PMID:21358755 |
RGD:9495920 |
NCBI chr X:131,858,148...131,944,639
Ensembl chr X:131,859,088...131,944,282
|
|
| G
|
Smarca5
|
SNF2 related chromatin remodeling ATPase 5
|
|
ISO
|
|
RGD |
PMID:21358755 |
RGD:9495920 |
NCBI chr19:44,172,465...44,208,967
Ensembl chr19:44,175,733...44,208,957
|
|
|
|
| G
|
Actb
|
actin, beta
|
|
ISO
|
|
RGD |
PMID:21358755 |
RGD:9495920 |
NCBI chr12:16,776,664...16,779,634
Ensembl chr12:16,776,661...16,780,242
|
|
| G
|
Actl6a
|
actin-like 6A
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr 2:117,413,206...117,436,758
Ensembl chr 2:117,418,601...117,436,754
|
|
| G
|
Actl6b
|
actin-like 6B
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr12:24,761,660...24,778,193
Ensembl chr12:24,761,693...24,778,151
|
|
| G
|
Arid1a
|
AT-rich interaction domain 1A
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr 5:151,192,014...151,269,291
Ensembl chr 5:151,192,014...151,265,386
|
|
| G
|
Arid1b
|
AT-rich interaction domain 1B
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr 1:47,973,199...48,328,793
Ensembl chr 1:47,973,997...48,328,793
|
|
| G
|
Arid2
|
AT-rich interaction domain 2
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr 7:129,326,371...129,443,813
Ensembl chr 7:129,324,137...129,443,187
|
|
| G
|
Brd7
|
bromodomain containing 7
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr19:34,882,238...34,910,944
Ensembl chr19:34,882,304...34,910,944
|
|
| G
|
Brd9
|
bromodomain containing 9
|
|
ISO
|
|
RGD |
PMID:21358755 |
RGD:9495920 |
NCBI chr 1:31,158,546...31,185,812
Ensembl chr 1:31,158,546...31,186,125
|
|
| G
|
Dpf1
|
double PHD fingers 1
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr 1:93,729,683...93,743,425
Ensembl chr 1:93,726,970...93,743,425
|
|
| G
|
Dpf2
|
double PHD fingers 2
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr 1:212,612,986...212,628,376
Ensembl chr 1:212,612,986...212,628,289
|
|
| G
|
Dpf3
|
double PHD fingers 3
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr 6:108,577,792...108,856,724
Ensembl chr 6:108,577,183...108,856,798
|
|
| G
|
Pbrm1
|
polybromo 1
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr16:6,219,933...6,319,336
Ensembl chr16:6,219,917...6,319,336
|
|
| G
|
Phf10
|
PHD finger protein 10
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr 1:64,625,373...64,642,110
Ensembl chr 1:64,624,465...64,642,049
|
|
| G
|
Smarca2
|
SWI/SNF related BAF chromatin remodeling complex subunit ATPase 2
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr 1:233,617,277...233,784,908
Ensembl chr 1:233,617,366...233,784,869
|
|
| G
|
Smarca4
|
SWI/SNF related BAF chromatin remodeling complex subunit ATPase 4
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr 8:28,438,370...28,535,071
Ensembl chr 8:28,443,810...28,535,070
|
|
| G
|
Smarcb1
|
SWI/SNF related BAF chromatin remodeling complex subunit B1
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr20:12,740,105...12,763,054
Ensembl chr20:12,740,943...12,763,055
|
|
| G
|
Smarcc1
|
SWI/SNF related BAF chromatin remodeling complex subunit C1
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr 8:118,986,917...119,093,161
Ensembl chr 8:118,989,516...119,093,152
|
|
| G
|
Smarcc2
|
SWI/SNF related BAF chromatin remodeling complex subunit C2
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr 7:1,466,223...1,494,627
Ensembl chr 7:1,466,454...1,494,626
|
|
| G
|
Smarcd1
|
SWI/SNF related BAF chromatin remodeling complex subunit D1
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr 7:132,708,627...132,719,167
Ensembl chr 7:132,708,432...132,719,167
|
|
| G
|
Smarcd2
|
SWI/SNF related BAF chromatin remodeling complex subunit D2
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr10:91,693,542...91,702,408
Ensembl chr10:91,693,542...91,702,522
|
|
| G
|
Smarcd3
|
SWI/SNF related BAF chromatin remodeling complex subunit D3
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr 4:11,441,142...11,472,781
Ensembl chr 4:11,441,157...11,472,781
|
|
| G
|
Smarce1
|
SWI/SNF related BAF chromatin remodeling complex subunit E1
|
|
ISO
|
|
RGD |
PMID:23355908 PMID:21358755 |
RGD:8694154, RGD:9495920 |
NCBI chr10:84,655,468...84,678,259
Ensembl chr10:84,655,468...84,676,185
|
|
|
|
|
Pathway Gene Annotations |
References Associated with the chromatin remodeling pathway:
- Polo SE J Mol Biol. 2014 Jun 2. pii: S0022-2836(14)00275-7. doi: 10.1016/j.jmb.2014.05.025.
- Hargreaves DC and Crabtree GR, Cell Res. 2011 Mar;21(3):396-420. doi: 10.1038/cr.2011.32. Epub 2011 Mar 1.
- Watanabe S and Peterson CL, Cold Spring Harb Symp Quant Biol. 2010;75:35-42. doi: 10.1101/sqb.2010.75.063. Epub 2011 Apr 18.
- Basta J and Rauchman M, Transl Res. 2014 May 10. pii: S1931-5244(14)00166-2. doi: 10.1016/j.trsl.2014.05.003.
- Clapier CR and Cairns BR, Annu Rev Biochem. 2009;78:273-304. doi: 10.1146/annurev.biochem.77.062706.153223.
- Bartholomew B Annu Rev Biochem. 2014;83:671-96. doi: 10.1146/annurev-biochem-051810-093157. Epub 2014 Mar 5.
- Swygert SG and Peterson CL, Biochim Biophys Acta. 2014 Aug;1839(8):728-36. doi: 10.1016/j.bbagrm.2014.02.013. Epub 2014 Feb 28.
- Hohmann AF and Vakoc CR, Trends Genet. 2014 Aug;30(8):356-63. doi: 10.1016/j.tig.2014.05.001. Epub 2014 Jun 3.
- Helming KC, etal., Cancer Cell. 2014 Sep 8;26(3):309-317. doi: 10.1016/j.ccr.2014.07.018.
- Gerhold CB and Gasser SM, Trends Cell Biol. 2014 Jul 31. pii: S0962-8924(14)00101-9. doi: 10.1016/j.tcb.2014.06.004.
- Erdel F and Rippe K, FEBS J. 2011 Oct;278(19):3608-18. doi: 10.1111/j.1742-4658.2011.08282.x. Epub 2011 Sep 2.
- Bartholomew B Curr Opin Struct Biol. 2014 Feb;24:150-5. doi: 10.1016/j.sbi.2014.01.010. Epub 2014 Feb 19.
- Allen HF, etal., Cell Mol Life Sci. 2013 Oct;70(19):3513-24. doi: 10.1007/s00018-012-1256-2. Epub 2013 Jan 23.
- O'Shaughnessy A and Hendrich B, Biochem Soc Trans. 2013 Jun;41(3):777-82. doi: 10.1042/BST20130027.
- Kolla V, etal., Cancer Res. 2014 Feb 1;74(3):652-8. doi: 10.1158/0008-5472.CAN-13-3056. Epub 2014 Jan 13.
- Luger K, etal., Nat Rev Mol Cell Biol. 2012 Jun 22;13(7):436-47. doi: 10.1038/nrm3382.
- Mueller-Planitz F, etal., Nat Struct Mol Biol. 2013 Sep;20(9):1026-32. doi: 10.1038/nsmb.2648.
- Narlikar GJ, etal., Cell. 2013 Aug 1;154(3):490-503. doi: 10.1016/j.cell.2013.07.011.
|
|
Ontology Path Diagram:
|
Import into Pathway Studio:
|
|