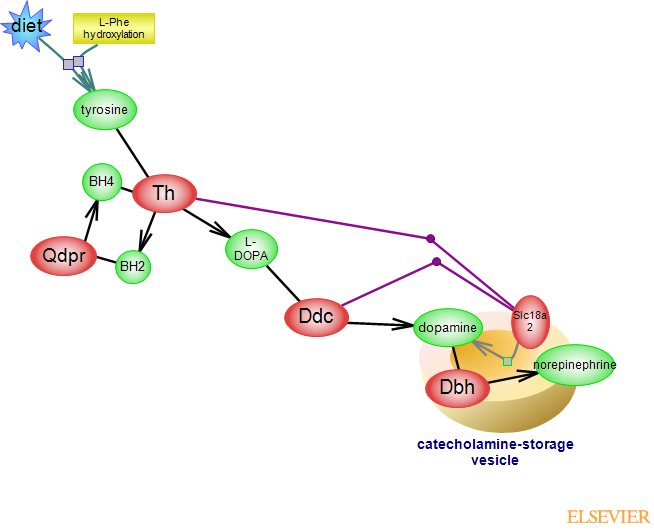
The synthesis and release of catecholamines dopamine, norepinephrine and epinephrine are triggered by various but distinct conditions and stresses. Norepinephrine and epinephrine, also known as noradrenaline and adrenaline, act as neurotransmitters and hormones. They signal via various adrenergic receptors, G protein-coupled receptors (GPCR) which couple to distinct G alpha subunits of heterotrimeric G proteins. Stresses such as cold and pain prompt the synthesis of norepinpehrine in sympathetic
nerve terminals and also adrenal medulla; hypotension or hypoglycemia, shock or emotional distress, prompt the synthesis of epinephrine, primarily in adrenal chromaffin cells. Dopamine ¿ the other catecholamine, whose synthesis takes place in cholinergic neurons, is a precursor for both norepinephrine and epinephrine but its roles are distinct. Catecholamine synthesis and the activities of participating enzymes are regulated by neuronal and humoral mechanisms. The precursor of all catecholamines is L-tyrosine which can be derived from the diet or from the hydroxylation of L-phenylalanine. Tyrosine hydroxylase (Th), an iron-containing, biopterin-dependent soluble enzyme catalyzes the conversion of tyrosine to dihydroxy-L-phenylalanine (L-DOPA) in the rate-limiting step of the reaction. The cofactor tetrahydrobioterin (BH4) is used in the reaction and the resulting dihydrobiopterin (BH2) is reconverted to BH4 by dihydropteridine reductase. Tetrahydrobiopterin can also be synthesized de novo from the purine nucleotide GTP. Th expression and activity is regulated in manifold ways. The short term control of Th activity is due to product(s)-based inhibitory feedback and activation by phosphorylation; several kinases and phosphatases have been implicated in modulating the activity of the enzyme. Long term control involves regulation of transcription and epigenetic mechanisms, stability and in humans, alternative splicing giving rise to four different isoforms. Th exists as a tetramer. Each monomer consists of a regulatory N-terminal stretch followed by the catalytic and tetramerization domains. Access the structure of the catalytic domain; or both catalytic and tetramerization domains. L-DOPA is converted to dopamine (DA) by the pyridoxal phosphate-dependent aromatic L-amino acid decarboxylase (Ddc). Both Th and Ddc interact with vesicle protein Slc18a2, which couples DA synthesis with transport into vesicles. Cytosolic DA is prone to oxidation to generate toxic quinones; the low pH inside the vesicle protects DA from oxidation. DA can be converted to norepinephrine which in turn, can be methylated to yield epinephrine. DA stored in vesicles is converted to norepinephrine by Dbh - a copper- and ascorbate-dependent enzyme that is found either in soluble or as a membrane-bound form within storage vesicles. To see a structure containing a fragment of this enzyme, click here.The soluble Pnmt converts norepinephrine to epinephrine using S-adenosyl-methionine (SAM) as a cofactor. To see the ontology report for annotations, GViewer and download, click here...(less)