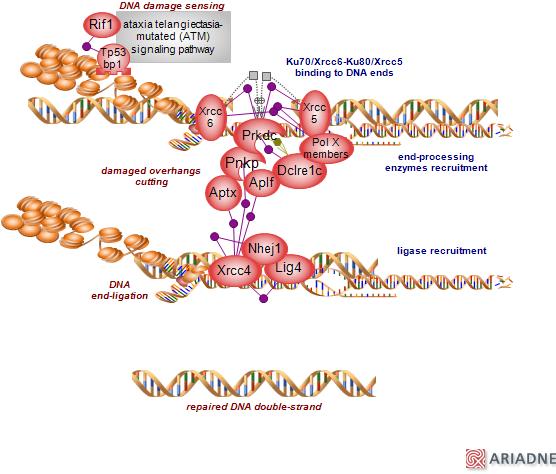
DNA lesions, particularly double-strand breaks (DSBs), can have severe genotoxic effects if not promptly handled. DSBs can be initiated by DNA damaging agents such as ionizing radiation or arise during replication through exposure to metabolites and are also physiological intermediates generated during V(D)J [variable (V), diversity (D) and the joining (J) recombination] and class switch recombination in lymphocytes of the immune system. The cells possess a robust response mechanism to DNA damag
e which triggers a range of signals to adequately handle the damage and maintain genomic integrity, collectively known as the DNA damage response (DDR). Central to mediating the response are the DDR kinases - ataxia telangiectasia-mutated (ATM), ATM and RAD3-related (ATR) and DNA-dependent protein kinase (DNA-PKcs). Of these, ATM plays a pivotal role - its signaling is activated by DSBs to promote the double-strand repair and prompt cell cycle checkpoint pathways, regulate cell fate decision (apoptosis or senescence), transcription and metabolic pathways. ATM, like the other DDR kinases, belongs to the phosphatidylinositol (PI) 3-kinase-like kinases (PIKK) family. It is a large serine/threonine kinase with an impressive repertoire of substrates, the size of which underlies the range of responses it can promote. As the name suggests, the human gene is mutated in the autosomal recessive condition associated with neurodegenerative and immunodeficiency phenotypes. A major function of ATM signaling is to activate the double-strand repair pathways. There are two principal DSB pathways: the non-homologous end-joining (NHEJ) and the homologous recombination (HR). The error-prone NHEJ which can be active throughout the cell cycle is the predominant one whereas the error-free HR is restricted to the S and G2 phases when sister chromatids are available to provide the necessary template. ATM signaling proceeds through several and overlapping layers, accompanied by activating positive feedback and regulatory negative inhibition loops. Two downstream sensors - Tp53bp1 and Brca1, prompt the unfolding of NHEJ or HR, respectively. Tp53bp1 opposes the resection reaction that is central to HR repair type, thus promoting NHEJ repair while Brca1 antagonizes Tp53bp1 to achieve the opposite effect, thus promoting the HR repair. The mechanisms by which the two proteins oppose each other to initiate the specific downstream repair route are incompletely understood. Tp53bp1 acts as a scaffold for additional interactors such as Rif1; the interaction with Rif1 appears to be important to antagonize resection. Tb53bp1 chromatin binding is dependent upon its homo-oligomerization and the dual recognition of dimethylated lysine 20 of histone 4 and ubiquitinated lysine 15 of histone 2A. In the absence of DNA damage/signal the dimethylated histone 4 - a known histone epigenetic mark, is masked from Tp53bp1 interaction. NHEJ is initiated by the binding of Ku heterodimer to the DNA ends of the DSBs (Xrcc6, known as Ku70 and Xrcc5, known as Ku80), followed by the recruitment of DNA-dependent protein kinase catalytic subunit (Prkdc) and several end-processing and accessory factors that include Dclre1c, known as Artemis, polymerases, polynucleotide kinase 3'-phosphatase (Pnkc), aprataxin (Aptx) and associated Aplf factor. The Artemis-Prkdc complex has several nuclease activities that include 5' and 3' endonuclease activity and hairpin opening activity while Dclre1c alone appears to have 5' exonuclease activity; Prkdc phosphorylates itself and Artemis. Polymerases that participate in NHEJ in somatic cells are Polm and Poll and in early lymphoid cells Dntt, in the context of V(D)J recombination - all are members of the X family that bind to the DNA-Ku complexes via their BRCA1 C terminus (Brct) domain. Processing of the DNA ends prepares them for relegation by a ligase complex that consists of Xrcc4, ligase 4 (Lig4) and the Xrcc4-like factor Nhej1 (known as Xlf). Xrcc4 acts as a scaffold: it interacts with itself as well as with Lig4, Nhej1 and factors such as Pnkc, Aptx and Aplf. Pnkc, as the name suggests, is both a kinase and a phosphatase, Aplf has both endo- and exonuclease activities, Aptx may important for deadenylation of aborted ligation products. The DNA-Ku end complexes can each recruit the various downstream activities in any order; the rather linear depiction of the pathway in the diagram is for ease of following. Components of the pathway are regulated at many levels. To access the ontology report for annotations, GViewer and download, click here...(less)